コロナワクチンはなぜ集団免疫の形成に失敗したのか(2)
 Googleの優先サイトに追加
Googleの優先サイトに追加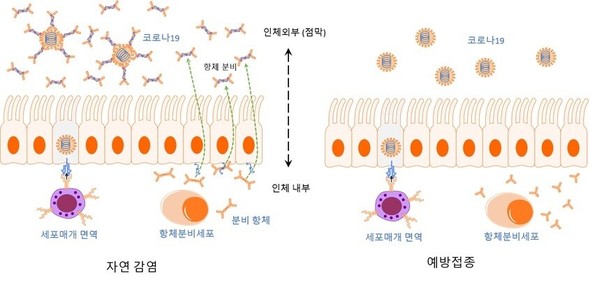
自然感染と予防接種免疫の違い
しかし理論と現実は異なる。ワクチンは集団免疫効果の達成に失敗した。何が原因なのだろうか。それはワクチンを投与する位置だ。図1は自然感染によって誘導される免疫(左)と、予防接種によるそれ(右)との違いを示している。前述のとおり粘膜は人体の外部であるため、粘膜環境に特化した抗体が分泌されウイルスを中和する。精巧で効率的な我々の免疫は、感染が起こった位置に防御力を集中させる。粘膜で感染が起きれば粘膜に抗体が分泌され、人体内部で感染が起きれば内部で抗体が作られる。図で描かれ方が異なっているように、粘膜と内部の抗体は文字通りクラスが異なる。だが、現在商用化されているすべてのコロナワクチンは、筋肉注射を通じて投与される。粘膜免疫の進行過程をすっ飛ばして、人体の内部に抗原を直に入れるのだ。この場合、人体の内部を循環する抗体は効率的に作られるが、粘膜抗体は分泌されない(正確には、ワクチンの種類によって結果は異なる)。コロナ感染は呼吸器の粘膜から始まるため、予防接種を受けたかどうかに関係なく、図の右側のように抗体が準備されていない状況になってしまうのだ。
幸い、このようなワクチンの死角地帯は長くは保たれない。感染が進めば粘膜の下の上皮細胞も死にはじめて外部との境界が破壊され、血管を流れる抗体が介入を開始するからだ。また、下で説明する細胞性免疫が介入し、危険な状況へと陥っていくのを遮断する。このような理由から、突破感染(ブレイクスルー感染)という用語は誤解の余地がある。コロナに自然感染して粘膜に分泌抗体ができている人でなければ、予防接種を受けたかどうかに関係なく、ひとまず感染は無条件に起きる。したがってワクチン接種後の突破感染は、実際の問題というよりは、PCRという敏感度の高い検査法のせいで検出される確率が高まったことを反映しているケースがほとんどだ。
だがこれとは別に、コロナはその短い間に増殖して広がる確率が高い。感染初期にできるだけ炎症を起こさずに密かに増殖する能力をコウモリの体内で育んだ後に、人のもとへとやって来たからだ。これこそ、新型コロナがパンデミックを引き起こした決定的な原因でもある。このようなコロナの固有の特性のため、集団免疫効果を得るために高いワクチン接種率が求められたのだ。
ウイルス工場を破壊する細胞性免疫
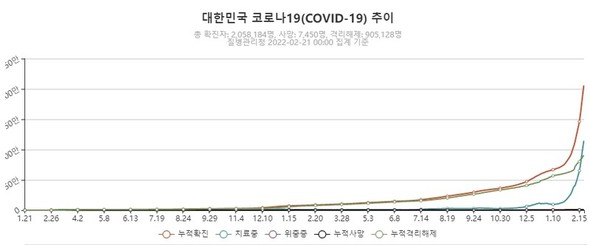
現実においてワクチン接種は本格化したものの、各国の接種率には大きな差が出はじめた。強い感染力を持つコロナと長期化するワクチン接種の組み合わせは最悪だった。そしてこの組み合わせは、集団免疫を完全に無力化するオミクロン株を誕生させた。ウイルスの変異を最小化する唯一の方法は、世界中で起こる増殖の絶対量を減らすことだ。簡単に言えば、感染者の数を抑えなければならないということだ。科学はそれを可能にするワクチンという道具を提供したが、国家主義は間違った使い方をした。変異を抑制するためには、すべての国ができるだけ短期間の内に同時に接種を進めなければならない。前のコラムで説明したように、ワクチンもコロナの進化を触発する選択圧だ。選択圧は短く、そして強く加えられないと適応変異が生じる。しかし、一部の国は3回目の追加接種(ブースターショット)を考えているのに、もっと多くの国では1回目の基本接種も遅々として進んでいないというのが現実だった。結局、専門家たちが懸念したとおり、ワクチンの死角地帯でオミクロン株が登場してしまったのだ。
オミクロン株の拡散は、デルタ株以前のパンデミックとは全く状況が異なる。デルタ以前の変異株は、抗体が認識するスパイク構造の変化が限られていたため、ワクチンを通じて作られる抗体の一部によって認知される確率がある程度あった。しかしオミクロン株はスパイクの構造が大幅に変わっているため、従来の抗体(ワクチンを通じてであれ、自然感染を通じてであれ)は全く認識できない。すなわち、ワクチンの投与位置の問題ではなく、抗体の観点から見て完全な新種のウイルスであるわけだ。そのため従来のワクチンの無用論が出ており、オミクロンスパイクで新たに設計されたワクチンでなければ接種する理由などないという主張もなされている。しかし、それらは私たちの体の免疫を単純に考え過ぎている。
普通、免疫といえばYの字型の抗体を思い浮かべ、それが免疫のすべてだと考えがちだが、抗体は複雑な免疫システムの一部に過ぎない。オミクロン株の登場によりワクチンによる集団免疫効果は無力化したが、個人の保護という予防接種の本質は依然として有効だ。なぜなら、抗体とともにウイルスに対抗する免疫の中心軸をなす細胞性免疫が健在だからだ。図1に示されているように、抗体がコロナのスパイクに結合して新たに細胞に感染するのを防ぐ中和作用をするとすれば、細胞性免疫はコロナに感染した細胞を殺す役割を果たす。もし細胞性免疫がなかったなら、ウイルス感染を解決するには膨大な量の抗体が必要になるだろう。ウイルスを新たに作り出す工場が稼動し続けているからだ。この工場をなくさなければ、抗体とウイルスは終わりなきイタチごっこを繰り広げなければならない。細胞性免疫はウイルスを作る細胞を殺す。すなわち工場を爆破する。したがって、予防接種者は重症化リスクが低くなる。
抗体が無力化しても細胞性免疫が2次防壁に
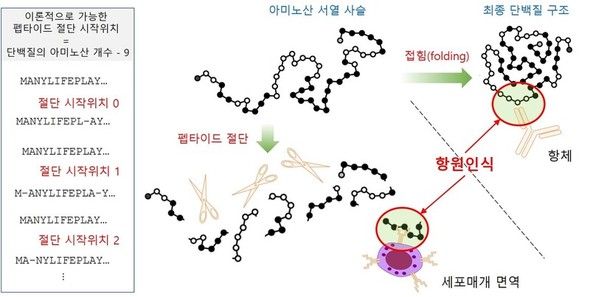
ワクチンが提供する同一のスパイク抗原によって刺激されているのに、なぜ抗体が無力化されても細胞性免疫は有効なのだろうか。その答えは図2で説明されている抗原の認識方式の違いにある。抗体はスパイクタンパク質の3次元構造を抗原として認識するのに対し、細胞性免疫はスパイクタンパク質が10あまりのアミノ酸単位に細かく刻まれた欠片、すなわちペプチドを抗原と認識する。つまり抗体は3次元構造を認識し、細胞性免疫は配列を認識する。この違いは抗原認識のレパートリーの違いを作る。どういうことなのか、スパイクタンパク質を例に説明してみよう。
現在商用化されているワクチンは、全てが武漢で初めて確認されたコロナウイルスのスパイクタンパク質を抗原として免疫に提示する。しかしコロナが変異し続けたことでスパイク構造の違いは少しずつ広がり、オミクロンに至ってはそれがかなり大きくなっている。抗体はスパイクの外部(図2の右側の白いアミノ酸の部位)のみを認識できる。スパイクタンパク質を構成する1274個のアミノ酸のうち、構造上折れ曲がった時に外部に突出しているアミノ酸だけが認識対象になるのだ。従って、この部位のアミノ酸が変わっただけでも、ワクチンによって作られた抗体は認識が困難になる。しかし、スパイクを構成する1274個のアミノ酸を十数個の大きさに切り刻むと、1264個のペプチドができる。スパイクタンパク質がいくら大きく変異していても、このペプチドすべてに変化が起きることはあり得ない。特にスパイクタンパク質の内部の中心をなす配列(図2の黒いアミノ酸の部位)は変異に弱い。タンパク質が折れ曲がる時、内部のアミノ酸が影響を与え合いながら構造の骨組みを作るからだ。ここに変異が発生すれば、そもそもまともな構造はできない。
まとめると、抗体が認識できない変異でも、細胞性免疫はウイルスのタンパク質によって認識できるということだ。このような理由から、集団免疫の成否如何にかかわらず、ワクチンによる免疫は重症化を防ぐ役割を依然として忠実に果たしているのだ。
訳D.K

